
肝X受体的基因多态性与代谢类疾病的相关性
赵慧佳1,李玲1,魏婉慧1,徐千1,时玉颖1,乐江1,王彦峰1,叶啟发1,2
1武汉大学中南医院,武汉大学肝胆疾病研究院,武汉大学移植医学中心,移植医学技术湖北省重点实验室,430071武汉,湖北;2中南大学湘雅三医院,卫生部移植医学工程技术研究中心,410013长沙,湖南
赵慧佳,女,大学本科,研究方向:基因多态性在器官移植中的应用。
叶啟发,男,通信作者,教授,主任医师,研究方向:肝胆外科,器官移植。E-mail:yqf_china@163.comTel:
摘要肝X受体(liverXreceptor,LXR)是人体内一种重要的核受体,为配体依赖性蛋白。LXR作为体内激素和营养信号的整合者,主要通过调控脂质和糖类等途径调节机体新陈代谢,参与Ⅱ型糖尿病、肥胖、心血管疾病、高脂血症等多种常见代谢疾病的发生发展。有研究证明,多个LXR基因多态性位点与代谢类疾病发生发展密切相关。研究LXR参与代谢性疾病的病理机制、探究LXR基因多态性与代谢性疾病的相关性,有利于从基因层面检测和评估代谢类疾病的发生风险,从而实现早期发现和预防该疾病的发生,提高代谢性疾病患者的生存质量,延长生存期。本文拟就近五年LXR参与代谢途径和其基因多态性与代谢性疾病的相关性做一综述。
关键词肝X受体;代谢性疾病;基因多态性
代谢类疾病(metabolicdisease)是当今社会最常见的一大类危害人体健康的慢性疾病的总称,包括Ⅱ型糖尿病、高脂血症、心血管疾病、肥胖等。目前,饮食控制和服用抗代谢疾病药物是治疗代谢类疾病患者的常规手段。然而,代谢类疾病成因复杂和不宜痊愈的特点极大限制了该类疾病的治疗效果。对于代谢类疾病的提前诊断和预防是临床上针对该类疾病的重要策略。肝X受体(liverXreceptor,LXR)作为一种人体内的重要核受体,参与多种代谢途径的调节,与代谢类疾病的发生密切相关。最新报道显示,LXR基因多态性与代谢类疾病发生风险密切相关,如果我们能够基于该方面的研究而提前采取针对性措施,就能有效预防该类疾病的发生发展[1]。由此,我们认为LXR有望成为代谢类疾病诊断和提前预测的新靶点。
1
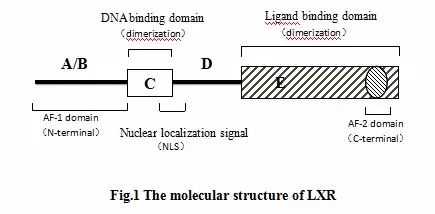
LXR的结构模式
LXR是人体内主要的核受体之一,为一种配体依赖性蛋白。LXR分为两种亚型----LXRα和LXRβ。LXRα又称为NR1H3,主要在脂质代谢旺盛的组织中表达,如肝细胞,脂肪细胞和巨噬细胞等。LXRβ又名NR1H2,在人体各种细胞中都有表达[2]。LXR作为配体激活的核受体,其主要以胆固醇作为配体而被激活。此外,胆固醇衍生物如氧甾酮和(24S)-24,25-环氧胆固醇等及LXR激动剂如GW3965和T0901317等物质都能作为配体激活LXR[3]。
LXR是细胞内的配体激活的转录因子,其与配体信号分子直接结合,控制转录应答和代谢过程。LXR由A、B、C、D、E五个区域组成,N-端为A/B区域,其包含一可变的配体非依赖性的转录激活域AF-1;C区域为高度保守的DNA结合域(DNAbindingdomain,DBD);D区域为非保守的铰链区,C、D区域之间存在核定位信号(nuclearlocalizationsignal,NLS);C-端为E区域----配体结合域(ligandbindingdomain,LBD),其包含一配体依赖的转录激活域AF-2。其中DBD与LBD分别介导DNA和配体与LXR的结合和二聚化的过程[4](见)。
在激活LXR的过程中,LXR与视黄醛X受体(retinoidXrecetor,RXR)结合形成异质二聚体,故LXR也可被RXR配体激活,如9-顺式维甲酸[5]。形成异质二聚体再与位于靶基因调控区的LXR
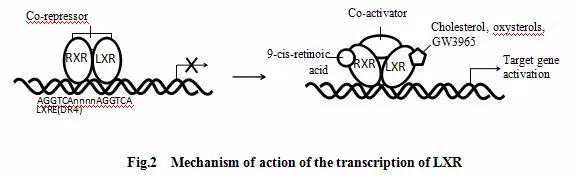
响应原件(LXRresponseelements,LXREs)相互作用。异质二聚体最初与辅助抑制因子结合,当其与LXRE(包含重复核苷酸序列AGGTCA,每次重复序列间隔四个核苷酸(DR4))相互作用后,辅助抑制因子被释放,辅助激活因子聚集,从而导致靶基因的转录[6](见)。
2
LXR参与的代谢过程
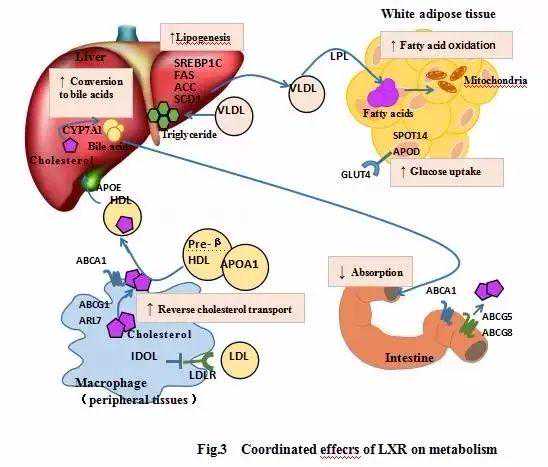
正常情况下,LXR主要通过参与胆固醇的逆向运输、促进胆固醇在肝脏中的代谢、促进胆固醇在小肠中的排泄并阻止其外周细胞的摄取等途径,发挥降低胆固醇、调节血脂的作用,在高胆固醇血症和动脉粥样硬化等疾病的发生发展中扮演着重要角色。另外,LXR与葡萄糖在脂肪细胞中的摄取和减少脂肪细胞葡萄糖氧化也有密切关系,间接发挥降低血糖的作用,具体作用机制见。
2.1胆固醇的逆向运输Venkateswaran[7]等首次证明,ATP结合盒转运蛋白A1(ATP-bindingcassettetransporterA1,ABCA1)是LXR的靶基因,受到LXR的高度调控。LXR主要通过ABCA1或其他转运体将细胞内的胆固醇转移到血浆的乏脂分子如载脂蛋白AI(apolipoproteinAI,APOAI)和高密度脂蛋白(highdensitylipoprotein,HDL)中,实现胆固醇的逆向运输。Fang[8-9]等人指出,ATP结合盒转运蛋白G1(ATPbindingcassettetransporterG1,ABCG1)作为LXR的另一靶基因,可使细胞内的胆固醇外流到HDL中,促使细胞内胆固醇逆向运输到肝脏形成胆汁酸。另有研究发现[10],LXR依赖诱导的胞内转运蛋白ADP核糖基化因子7(ADP-ribosylationfactor-like7,ARL7)亦可以促进胞内胆固醇流向胞膜,从而诱导胆固醇的外流。此外,LXR还可通过一增强子区域诱导一组载脂蛋白(APOE,APOC1,APOC2和APOC4)的表达,进而调节脂质的运输和分解代谢[11]。
2.2胆固醇在肝脏中的代谢由于胆固醇不能在动物细胞中分解代谢,所以胆固醇必须逆向运输到肝脏转化为胆汁酸或随胆汁酸排泄。小鼠细胞色素P4507A1(cytochromeP4507A1,CYP7A1)为胆固醇转化为脂肪酸的限速酶。Peet[12]等构建LXRα基因敲除小鼠模型发现,该小鼠进行高胆固醇饮食后,肝脏中积累大量胆固醇,且进行高脂饮食后正常小鼠CYP7A1的mRNA表达量显著增加,而基因敲除小鼠的mRNA表达量并无变化,从而首次证实了CYP7A1为LXR的靶点。另有研究报道,LXR可通过诱导固醇调节元件结合蛋白-1c(sterol-regulatoryelement-bindingprotein1C,SREBP1C),乙酰辅酶A羧化酶(acetylCoAcarboxylase,ACC),固醇酰辅酶A脱氢酶(sterolCoAdesaturase,SCD1),脂肪酸合成酶(fattyacidsynthase,FAS)等蛋白质的合成进而促进脂肪的生物合成,从而为游离胆固醇的酯化提供基板,促进游离脂肪酸的酯化,减少游离胆固醇对肝脏的毒性[13-14]。
2.3胆固醇在小肠中的排泄和外周细胞的摄取Kawase[15]等研究发现,LXR通过诱导小肠吸收细胞上的转运体ATP结合盒转运蛋白5(ATP-bindingcassettesub-familyGmember5,ABCG5)和8(ABCG8)的表达,进而促进小肠吸收细胞内胆固醇的外流,减少体内胆固醇的含量。刺激LXR进而诱导ABCG5的表达,使小肠排泄胆固醇的量增加一倍[16]。在外周细胞中,LXR和固醇调节元件结合蛋白(sterolregulatoryelement-bindingprotein,SREBP)互为拮抗分子,共同维持胆固醇代谢的平衡,当细胞内胆固醇水平较低时,SREBP2作用于低密度脂蛋白受体(Lowdensitylipoproteinreceptor,LDLR),使细胞摄取更多胆固醇[17],而当细胞内胆固醇水平过高时,LXR被激活,并诱导可诱导的LDLR降解器(inducibledegraderofLDLR,IDOL)----E3连接酶作用于LDLR,使其泛素化和酶解,从而使LDLR失活,抑制胆固醇的摄取[18]。
2.4葡萄糖的代谢有报道表明,胰岛素可以诱导激活LXR,使其与LXR配体葡萄糖结合,编码激活LXR的下游靶点葡萄糖转运体(glucosetransporter-4,GLUT4),然而,由于LXR还可以抑制许多葡萄糖摄取的相关蛋白的表达,所以LXR对葡萄糖的总体效果是抑制葡萄糖在脂肪细胞中的摄取[19-22][23]。
3
LXR的基因多态性与代谢相关疾病的关系
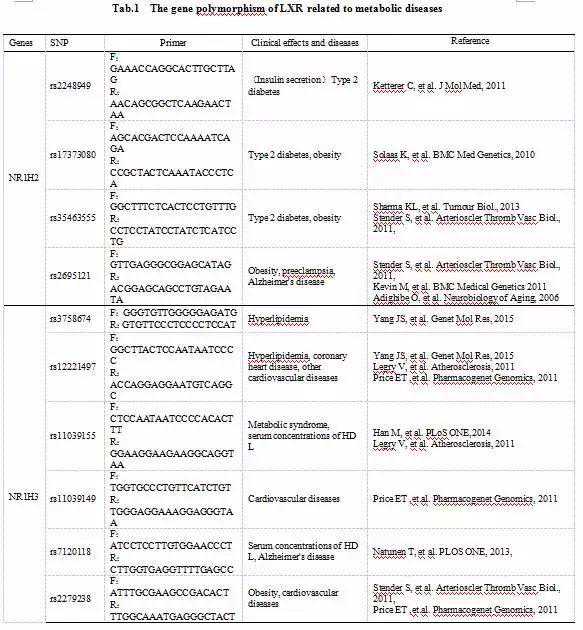
LXR的基因多态性均与代谢类疾病(包括Ⅱ型糖尿病,肥胖,阿尔兹海默综合征等)密切相关(见)。其中,LXRα位于染色体11上,由17个外显子组成,LXRβ,位于染色体19上,由10个外显子组成,两种亚型所相关的代谢性疾病略有不同。
3.1Ⅱ型糖尿病与肥胖Ketterer[24]等证明,多个LXRβ单核苷酸多态性(singlenucleotidepolymorphism,SNP)与胰岛素的分泌有关。在该项研究中,作者对欧洲糖尿病患者和非糖尿病患者共1574人进行调查,通过基因测序收集了6个SNP位点。结果发现,位于NR1H2基因上的rs2248949与第一时相胰岛素分泌相关,T等位基因携带者的胰岛素分泌量比C等位基因纯合个体减少6%;此外,该基因位点突变还影响胰岛素原转换为胰岛素过程,T等位基因携带者的胰岛素产生量比C等位基因纯合个体减少18%。由此,我们推测,rs2248949影响胰岛B细胞功能,参与Ⅱ型糖尿病的发生发展。
有研究报道[25],NR1H2基因rs17373080与挪威人群Ⅱ型糖尿病及欧洲人群的肥胖密切相关,该SNP的G等位基因携带者中BMI大于30的人数要比其他实验者中BMI大于30的人数增长59%。Dahlman[26]等发现,rs17373080C等位基因携带者的LXR转录活性更高,与Ⅱ型糖尿病的发病机制有着密切关系,从而使不患Ⅱ型糖尿病的人中含C等位基因的人数大于不含C等位基因的人数,且具有统计学意义。此外,NR1H2基因的rs35463555同样也被报道和Ⅱ型糖尿病的发展有关,不同等位基因的患病人数之间差异具有统计学意义。
另有瑞典的研究者发现[27],NR1H3基因的rs2279238的多态性使得LXRα在脂肪细胞中的表达量不同,从而导致调查人群中肥胖与该SNP的关系差异具有统计学意义,由此研究作者还证明了肥胖女性的LXRα的mRNA的表达水平较高。该文章还证实了NR1H2基因的rs35463555、rs2695121的多态性影响着肥胖的发生发展,肥胖人群和非肥胖人群的基因型差别具有统计学意义。
3.2高血脂Robitaille[28]等人在对35名法国高血脂和高LDL浓度的患者进行LXRα的基因分型时发现,-115A(rs12221497)、-840A(rs61896015)或-1830C(rs3758674)三种等位基因携带者的总血脂浓度要高于-115G/G、-840C/C或-1830T/T纯合子的患者。上述三种SNP的A或C等位基因携带者在摄入高脂饮食后更容易引起血脂水平升高。因此,我们推测,胆固醇的摄入和LXRα多态性的协同作用,引起血脂变化,进而可能导致心血管疾病的发生。
Legry[29]等收集了2290个法国患者临床病例,发现NR1H3基因的rs11039155的SNP与代谢综合征有关。携带A等位基因的个体可以降低30%的患代谢综合征的风险,同时携带A等位基因的个体与血浆高密度脂蛋白胆固醇(HDL-C)浓度升高有关联,从而可能减少患Ⅱ型糖尿病和心血管疾病的风险。
VanessaLegry[30]等在对欧洲1144个青少年LXRα基因的SNP的研究中发现,rs11039155的A等位基因与其下游的几个靶基因(ABCA1,CETP,LPL)的SNP的次要等位基因连锁,与血清高HDL-C浓度相关;而rs12221497的A等位基因与其下游靶基因(ABCA1,APOEε4)的SNP的次要等位基因连锁,与血清低HDL-C浓度相关。由此我们认为,LXRα和其下游靶基因的基因多态性导致了青少年有关HDL-C浓度的遗传变异。
3.3代谢相关的心血管疾病
Elvin[31]等人从INVENT-GENES研究基础上,根据性别和种族的不同选取297个个体,对LXRα的7个SNP位点进行研究,观察全因死亡率、非致死心肌梗死和非致死性中风等实验室指标。证明在非黑人种族中有三个SNP与该主要指标相关,NR1H3基因上的rs11039149的G等位基因和rs1222497的A等位基因能够减少发生三项主要指标的风险,而rs2279238的T等位基因则是增加发生主要指标的风险。且rs2279238的T等位基因的携带者其治疗方法与基因型之间有显著相关性,由此论证了LXRα参与多种心血管疾病的病理过程,其遗传信息改变将影响该疾病的发生发展风险。
Zhou[32]等人对中国汉族人口进行的调查研究中发现,NR1H3的rsl2221497基因多态性与冠心病的发生相关,该位点基因型为AA和GA的冠心病患者人数是基因型为GG患者的1.76倍,而且A等位基因携带者比G等位基因携带者患冠心病的风险增加0.88倍。通过Logistic回归分析并排除其他次要因素后,证明rs12221497的A等位基因可作为患冠心病的风险基因。
某研究小组[27]在对61710个丹麦白种人进行缺血性血管疾病与基因多态性的研究中发现,NR1H3的两个SNP连锁-840AA/-115AA(rs12221497)的个体在预测缺血性心脏病(ischemicheartdisease,IHD)风险比率达1.2,在预测心肌梗死的风险比率中达1.5。其中,10281个丹麦白种人缺血性脑血管病的发病风险中达1.7。1830TC(rs3758674)与上述两个基因连锁所得的单倍型(1830C/840A/115A)与LXRα的转录紧密相关。
3.4阿尔兹海默综合征有研究表明[33],NR1H3的rs2695121的C/C基因型与ApoE-ε4的出现相关,而ApoE-ε4是迟发型阿尔兹海默综合征(Alzheimer’sdisease,AD)发生的变异风险因素。当血红素加氧酶(hemeoxygenase,HO-1)基因的rs2071746的T/T基因型与LXRβ基因的rs2695121的T/T基因型、rs1052533的A/A基因型、rs1405655的T/T基因型中的任何一个相关联,将会导致HO-1和LXRβ的双重低表达,从而导致胆固醇的堆积,Aβ产物的合成增加,从而导致发生AD的风险增加[34]。另有研究报道[35],NR1H3基因的rs7120118的C/C基因型比T/T基因型患者可溶性Aβ42水平要低,从而降低患AD的风险。该SNP的C/T基因型比T/T基因型患者合成mRNA的水平要高,NR1H3的SNP会影响LXRα的表达和可溶性Aβ42的水平。
4
小结与展望
LXR作为一种在人体各种细胞中均表达的核受体,它对机体代谢过程的调节作用不可忽视,LXR通过对其下游参与机体代谢的基因(如ABCA1,SREBP-1C,GLUT4等)的调控进而对机体的新陈代谢产生影响。而LXR的基因多态性将会直接影响到其下游基因的表达,从而使不同人对代谢性疾病的易感性不同。因此,我们旨在对LXR的重点SNP位点(如近起始密码子位点rs35463555等)与代谢性疾病的发生关系进行进一步的详细研究,希望能根据该研究为临床代谢疾病的诊断和防治提供新的思路。
此外,LXR多态性的研究不仅局限于原发性代谢性疾病,近期文献报道,由于供受体状态、激素及免疫抑制剂等原因,移植患者术后极易并发肥胖、高血压、糖尿病和代谢综合征等代谢类疾病[36-38]。尽管前期,Kiss[39]等已经初步证实,LXR的活性与肾移植术后移植物的损伤具有相关性,且有报道表示,LXR与干细胞移植术后重要器官的损伤有着密切的关系[40-41]。但LXR在移植领域研究和LXR基因多态性与其术后并发症的关系还处于初步探索阶段。由此,我们推测,LXR参与的代谢途径和其基因多态性与代谢类疾病的关系,与患者术后继发代谢类疾病的发生存在相关性,其同样有进一步研究的必要。
目前,针对LXR基因多态性的研究已初有成效。然而,对LXR已发现的SNP位点的作用机理仍不清楚,而对LXR上下游基因多态性也没有深入研究。有报道称,LXR下游基因ABCA1、SREBP-1的多态性也与代谢类疾病相关[42-43],因此我们认为对LXR自身和其上下游基因的联合研究更有利于对其作用机理的探究和对代谢类疾病发生概率的全面筛查。
综上所述,临床上,以LXR及其上下游基因的基因位点作为靶点进行联合检测,可以较快、较准确的诊断出患代谢类疾病的风险,并加以预防,同时还可以根据基因靶点的多态性,提出不同的治疗方案,使用不同剂量的药物对疾病进行治疗,从而达到临床上的个体化治疗。因此,对LXR基因多态性的探究为原发以及术后继发的代谢类疾病的诊断、预防和治疗开辟了一条新途径。
参考文献
[1]RankinenT,SarzynskiMA,GhoshS,,cardiovasculardiseaseoutcomes,andcardiov-ascularriskfactors[J]?CircRes,2015,116(5):909-922.
[2]BiXK,SongJL,GaoJ,ucedIL-8expressioninothelialcellsviatheNF-κBpathwayandSUMOylation[J].JCellMolMed.,2016,20(10):1-10.
[3]ZhangL,ChenYL,YangXX,/2inhibitorsactivatemacrophageABCG1expressionandreversecholesteroltransport—Ananti-atherogenicfunctionofERK1/2inhibition[J].BiochimicaetBiophysicaActa,2016,(1861):1180-1191.
[4]KissM,CzimmererZ,NagyL,riticcellfunction:Fromphysiologytopathology[J].JAllergyClinImmunol,2013,132(2):264-286.
[5]HuwaitEA,SinghNN,sporterA1expressionbyliverXreceptor/retinoidXreceptoragonistinhumanmacrophages[J].JCellBiochem,2015,116(9):2032-2038.
[6]NagyL,SzantoA,SzatmariI,etheirlipidenvironmentandshapetheirimmuneresponse[J].PhysiolRev,2012,92(2):739-789.
[7]VenkateswaranA,LaffitteBA,JosephSB,LXRα[J].;97(22):12097-12102.
[8]HongYF,KimH,24(S),25-Epoxycholesterol-MediatedABCA1andABCG1ProductionandCholesterolEffluxinMacrophages[J].PLoSOne,2016,11(4):1-18.
[9]FangL,ChoiSH,BaekJS,[J].Nature,2013,498(7452):118-122.
[10]HongC,WalczakR,DhamkoH,flammatorygeneexpression:identificationofARL7asadirecttarget[J].JLipidRes,2011,52(3):531-539.
[11]KirchgessnerTG,SlephP,OstrowskiJ,teinmetabolismandcirculatingneutrophils[J].CellMetabolism,2016,24(2):223-233.
[12]PeetDJ,TurleySD,MaW,earoxysterolreceptorLXRα[J].Cell,1998,93(5):693-704.
[13]HsiehJ,KosekiM,MoluskyMM,tohepatitis[J].Nature,2016,535(7611):303-307.
[14]HøgmoenÅstrandOA,GiklingI,SylteI,elipogenesisinhumancellmodels[J].EurJMedChem,2014,74:258-263.
[15]KawaseA,HataS,TakagiM,andg8toinfluenceintestinalcholesterolabsorption[J].JPharmPharmSci,2015,18(5):765-772.
[16]vanderVeenJN,vanDijkTH,VrinsCL,nofplasmacholesterol[J].JBiolChem,2009,284(29):19211-19219.
[17]KartawijayaM,HanHW,KimY,2[J].FoodNutrRes,2016,60(31120):302-309.
[18]GuoD,ReinitzF,YoussefF,/AKT/SREBP-1/LDLR-depentpathway[J].CancerDiscov,2011,1(5):442-456.
[19]ShavvaVS,BogomolovaAM,NikitinAA,atomaCellLineHepG2:TheRoleofInteractionbetweenFOXO1andLXRβTranscriptionFactors[J].JCellBiochem,2016,doi:10.1002/
[20]MitroN,MakPA,VargasL,[J].Nature,2007,445(7124):219-223.
[21]PetterssonAM,StensonBM,Lorente-CebriánS,[J].Diabetologia,2013,56(9):2044-2054.
[22]Korach-AndréM,ArcherA,BarrosRP,(LXR)isoformscontrolenergyexpiturebyregulatingbrownadiposetissueactivity[J].ProcNatlAcadSciUSA,2011,108(1):403-408.
[23]StensonBM,RydénM,SteffensenKR,pocytes[J].ocrinology,2009,150(9):4104–4113.
[24]KettererC,MüssigK,MachicaoF,βassociateswithinsulinsecretioninsubjectsatincreasedriskfortype2diabetes[J].JMolMed,2011,(89):75-81.
[25]SolaasK,LegryV,RetterstolK,ismswithtype2diabetesmellitusandobesityinthreecohortstudies:HUNT2(Norway),MONICA(France)andHELENA(Europe)[J].BMCMedicalGenetics,2010,11(144):234-244.
[26]SharmaKL,MisraS,KumarA,(LXRs)geneticvariantstogallbladdercancersusceptibility[J].TumourBiol,2013,34(6):3959-3966.
[27]SterS,Frikke-SchmidtR,AnestisA,iseaseintheGeneralPopulation[J].ArteriosclerThrombVascBiol,2011,(31):2990-2996.
[28]YangJS,HaoJJ,WangSS,αpolymorphismandstrokeinaChinesepopulation[J].GenetMolRes,2015,14(1):1757-1762.
[29]HanM,LiangL,LiuLR,:EffectonSusceptibility[J].PLoSONE,2014,9(5)::10.1371/
[30]LegryV,BokorS,BeghinL,ralphaanditstargetgeneswiththeserumHDL-cholesterolconcentrationinadolescentsoftheHELENAStudy[J].Atherosclerosis,2011,(216):166-169.
[31]PriceET,PacanowskiMA,MartinMA,αgenepolymorphismsandvariablecardiovascularoutcomesinpatientstreatedwithantihypertensivetherapyresultsfromtheINVEST-GENESstudy[J].PharmacogenetGenomics,2011,21(6):333-340.
[32]ZhouYF,ZhangJ,LiZX,α(LXRα)genepolymorphismandcoronaryheartdisease,serumlipidsandglucoselevels[J].LipidsHealthDis,2014,13(34):25-29.
[33]AdighibeO,ArepalliS,DuckworthJ,(NR1H2)maycontributetotheriskofAlzheimer’sdisease[J].NeurobiolAging,2006,27(10):1431-1434.
[34]InfanteJ,Rodríguez-RodríguezE,MateoI,Alzheimer’sdiseaserisk[J].NeurobiolAging,2010,31(4):710-714.
[35]NatunenT,MartiskainenH,SarajärviT,ndtheprogressionofAlzheimer’sdisease[J].PLoSOne,2013,8(11):e80700.
[36]汪江林,左笑丛,杨梦等.钙调神经磷酸酶/NFATc信号通路介导他克莫司诱导移植后糖尿病的研究进展[J].中国临床药理学与治疗学,2014,19(1):74-80.
[37]SabineG,MaximilianJ,MaciejM,:ananalysisofdatafromfastedpatient.[J].Transplantation,2015,99(6):1243-1249.
[38]ChangAL,CortezAR,BondocA,:Apreoperativeandpostoperativeconcern[J].Surgery,2016,160(4):1111-1117.
[39]KissE,PopovicZ,BedkeJ,(LXR)activationrelevantcontributionofmacrophageLXRα[J].AmJPathol,2011,179(1):92-103.
[40]WangY,LiC,ChengK,enchymalStemCellstoAttenuateMyocardialIschemiaInjuryThroughTLR4/NF-jBandKeap-1/Nrf-2SignalingPathways[J].AntioxidRedoxSignal,2014,21(18):2543-2557.
[41]BhargavaM,VikenKJ,DeyS,ntation[J].BiolBloodMarrowTransplant,2016,22(8):1383-1390.
[42]ErgenHA,ZeybekU,GokO,mellitus[J].BiochemMed(Zagreb),2012,22(1):114-120.
[43]KalantarianS,RimmEB,HerringtonDM,,geneticvariation,andprogressionofcoronaryatherosclerosisamongwomen[J].AmHeartJ,2014,167(4):627-635.



